Joshua Weitz, from the University of Maryland, visited our lab last week. Joshua is a mathematical biologist who studies the ecology and evolution of viruses. He gave a fascinating departmental seminar about how viruses may evolve to have intermediate efficiency at exploiting their hosts, and the implications of this for managing disease outbreaks in human populations. He explores related ideas in his latest book Asymptomatic: The Silent Spread of COVID-19 and the Future of Pandemics, published by Johns Hopkins University Press.
Last week, Ryan travelled to the Helmholtz Centre for Environmental Research in Leipzig, Germany, to attend a workshop on tree community dynamics in species-rich tropical forests. The workshop brought together experts on tropical forests and related areas of ecology from around the world. Ryan gave two presentations, covering topics including our lab’s work on unified theories of biodiversity and on spatial synchrony in tropical tree communities.
Randall has completed his Honours project on species–area relationships for fish communities in freshwater lakes. Tying in with our lab’s previous work on mangrove tree communities, Randall was specifically interested in testing for a flat phase in the species–area relationships for small lake areas. Under a theoretical framework developed in our lab would be interpreted as a niche-structured phase that occurs when immigration is very low. While Randall found evidence of a flat phase in some of his ten datasets, it was not a pervasive phenomenon, which he speculated may be because fish are more effective at dispersing between lakes than we had supposed. Randall will be starting a PhD in the lab in August.
We recently hosted Professor Jedediah Brodie from the University of Montana. Jed’s work concerns global biodiversity conservation and he has a strong research focus in Borneo, where he has carried out extensive field work and conducted modelling to inform planning of protected areas and wildlife corridors. During his visit, Jed gave a departmental seminar and discussed potential research synergies with us.
Last week Professor Wang Shaopeng, from Peking University, visited us to meet with staff and students and to deliver a departmental seminar. Shaopeng is a theoretical ecologist who studies the relationship between biodiversity, ecosystem function and ecosystem stability. In his seminar, Shaopeng discussed, among other topics, his recent work on spatial scaling laws for biodiversity and ecosystem stability.
Our lab recently received a grant from Singapore’s Ministry of Education to build a new framework for biodiversity accounting. The global biodiversity crisis has spurred efforts to quantify the relative biodiversity value of patches of land under different scenarios. Biodiversity accounting can support biodiversity impact assessment, biodiversity footprint estimation, and biodiversity credit market development. The World Economic Forum projects that annual demand for biodiversity credits could reach US$2 billion by 2030 and US$69 billion by 2050. Unfortunately, current frameworks for biodiversity accounting are mostly ad hoc and lead to misleading valuations in some cases. The biodiversity accounting framework we will develop under this new grant will be based on fundamental ecological and economic principles and will ultimately support more scientifically sound allocation of economic resources to protect natural capital.
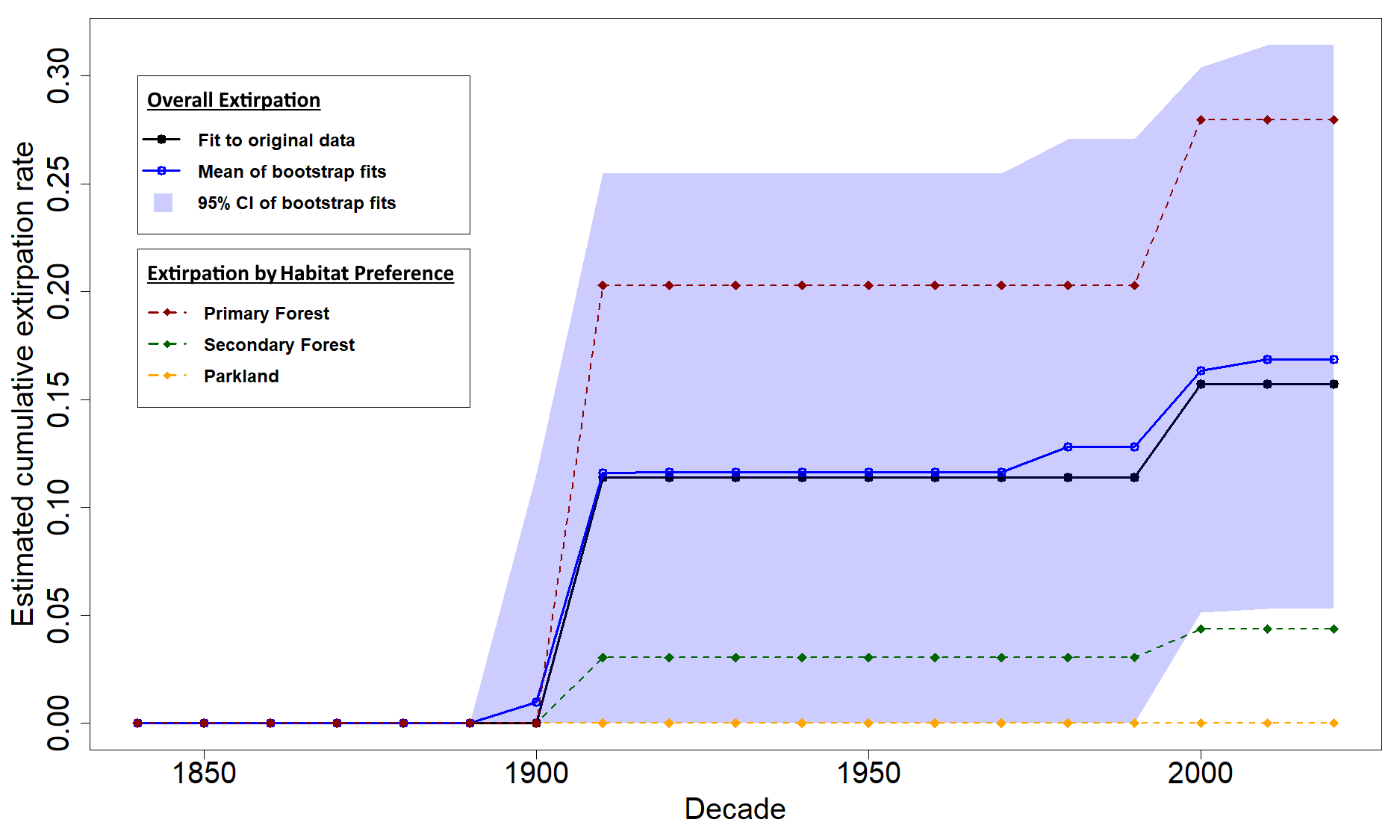
Singapore has experienced substantial habitat and biodiversity loss since 1819. Nevertheless, the last century has also seen some forest recovery, as well as the natural re-establishment of several previously extirpated species like oriental pied hornbills and smooth-coated otters. Animals that can fly and swim can re-establish from source populations in Malaysia, but snakes and lizards—also known as squamates—are not able to recolonise as easily. How can we use mathematical models to think about which species to reintroduce to recovering forests? In 2023, we published a paper applying the novel MODGEE (matrix-of-detections-gives-extinction-estimates) model to detailed historical bird, butterfly and plant datasets from Singapore to study extirpation rates for these groups. In a new study, led by Sankar and just published in Biological Conservation, we applied this model to squamates in Singapore.
We compiled a detailed historical dataset for squamates in Singapore (largely based on records from Figueroa et al., 2023) and used the MODGEE model first to generate an extirpation timeline. Our estimated extirpation rate for squamates was 17%, which was lower than previously estimated rates for birds (40%) and butterflies (46%). This implies that squamates may be more resilient to habitat degradation than birds, perhaps because birds generally have lower population densities, and butterflies, perhaps because of butterflies’ reliance on host plants.
Our model estimated that about 17% of squamates have been extirpated from Singapore over the last two centuries. Primary forest species (red) experienced higher extirpation rates than secondary forest-tolerant (green) and parkland species (yellow).
We also used the results to inform a conservation translocation triage for extirpated squamates in Singapore. Given that squamates were never systematically hunted at scale (no pun intended) in Singapore, their extirpation timeline can serve as an indicator of each species’ sensitivity to habitat loss. Since highly sensitive species would have been lost early on in Singapore’s developmental history, recently extirpated species would likely be better candidates for reintroduction to Singapore’s recovering forests. Through this method, we identified the hulk forest gecko (Gekko hulk) as a fitting potential candidate species for reintroduction to Singapore from stock populations in Peninsular Malaysia.
We identified the hulk forest gecko (Gekko hulk) as a candidate for reintroduction to Singapore.
The Biological Sciences Graduate Congress is an annual event bringing together Master’s and PhD students from NUS; University of Malaya, Malaysia; and Chulalongkorn University, Thailand. Ho Ning and Tricia recently attended the 2025 congress at Chulalongkorn University. Ho Ning presented her work on modelling sea turtle population dynamics in Southeast Asia. Tricia presented her work on estimating dark extinctions for freshwater taxa in Singapore.