Last week Ryan attended the OceanX summit in Singapore. OceanX is a nonprofit organisation dedicated to marine research and conservation. The summit was held aboard the organisation’s research vessel, the OceanXplorer. The summit brought together scientists, creative professionals, and philanthropists to brainstorm new ways to further ocean science and communicate it to the public.
Ryan has just returned from the Ecological Society of America’s 110th annual meeting in Baltimore, Maryland, USA. In addition to partaking of the stimulating smorgasbord of seminars, Ryan attended the awards ceremony to receive the George Mercer Award on behalf of himself and Lynette Loke, and he launched his new textbook. The textbook is titled Theoretical Ecology: Concepts and Models with R and is available from Princeton University Press in paperback and hardback.
The ability of a scientific model to make accurate predictions is an important criterion for assessing its validity, but in ecology there are relatively few studies that have made and tested true a priori predictions, i.e., predictions of unseen data. In a study led by Tak and just published in Ecology Letters, we tested several ecological models’ predictions of unseen higher-order diversity patterns in island archipelago data.
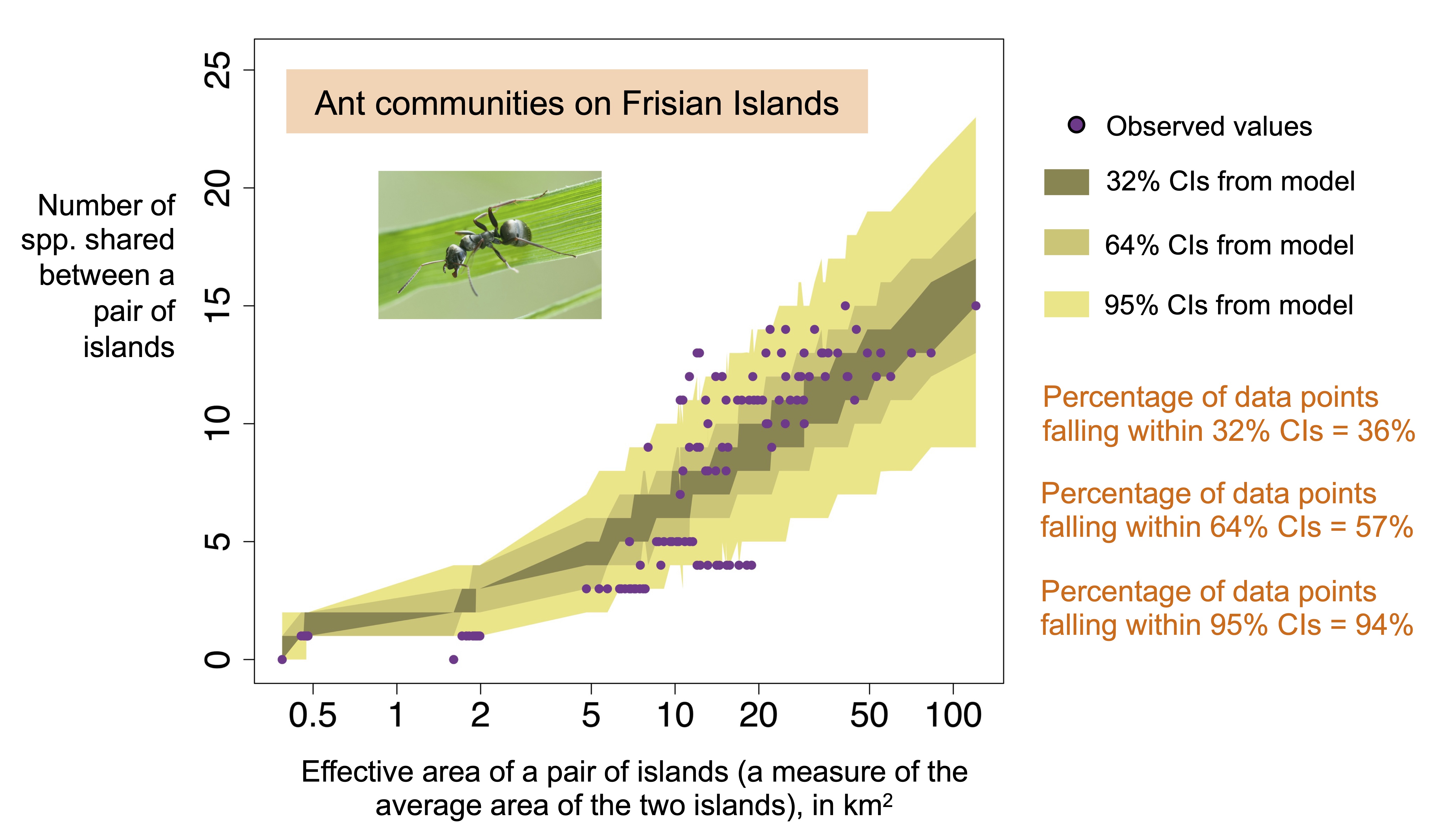
Specifically, we (i) fitted a suite of mechanistic models to observed values of alpha diversity (i.e., number of species on each island) for each of 17 archipelagos, (ii) used the fitted models to make quantitative predictions of three higher-order patterns of island biodiversity, and (iii) quantitatively tested the predictions. The 17 datasets represented a wide range of taxa (including plants, birds and mammals) and archipelago types (including marine and inland water) (Fig. 1). Importantly, the predicted patterns of biodiversity represent higher orders of diversity that contain information absent in alpha diversity, namely the number of species shared between each pair of islands, the number of species shared among each triplet of islands, and the occupancy–frequency distribution that shows the frequency distribution of the number of islands occupied by species in an archipelago.
Our central finding is that an individual-based neutral model of island community dynamics produced fairly good predictions of island biodiversity (e.g., Fig. 2). This suggests that stochastic neutral competition among species together with dispersal limitation is a parsimonious explanation for multiple patterns of island biodiversity. A non-neutral version of the model that included coarse niche structure had worse predictive ability, probably due to overfitting. Our study is a clarion call for further efforts to test true predictions in ecology, in particular using mechanistic models to shed light on the processes structuring ecological systems in nature.
Fig. 1. An aerial view of Baltrum, one of the Frisian Islands in the North Sea. The Frisian Islands form one of the 17 archipelagos that we examined. This photo was taken by Carsten Steger and used in an unaltered form under a CC BY-SA 4.0 license.Fig. 2. For ant communities on the Frisian Islands (17 islands), the observed relationship between number of species shared between each pair of islands and the average area of the islands (points) is accurately predicted by an individual-based neutral model fit only to the observed relationship between the number of species on each island and island area (this information by itself imposes only very weak constraints on the quantity being predicted). The proportion of points that fall within each confidence interval (shaded regions) closely tracks the nominal coverage of the interval (see text at right). The photo shows an ant of the species Formica fusca, which is found on the Frisian Islands. The photo was taken by Mathias Krumbholz and used in an unaltered form under a CC BY-SA 3.0 license.
The Ecological Society of America (ESA) has awarded Lynette and Ryan the George Mercer Award for their paper testing theories of community assembly using an experiment on intertidal communities in Singapore (Loke & Chisholm 2023). The George Mercer Award was established in 1948 and is given annually for an outstanding ecological research paper published in the last two years whose lead author is under 40. The ESA announcement wrote, “Deftly integrating field experiments with clear hypotheses and mathematical modeling, this study provides novel insights into an ecological question that has been debated for decades. The results of the paper advance our understanding of the forces structuring ecological communities, and the authors combine theory and empirical evidence in a creative and exceptionally well-written way.”
Lynette has collaborated with our lab on several projects in recent years. She moved to the University of Melbourne last year to start a spatial and community ecology lab.
Professor David Waxman, from Fudan University, just completed a two-week visit to our lab. David is a mathematical biologist who works on population genetics and evolution. During his visit, he engaged in productive and stimulating discussions with lab members, and he delivered a departmental seminar about a new multiallelic selection model he has been developing.
Co-operation is pervasive in human societies, but the evolutionary origins of co-operation are still incompletely understood. One prominent hypothesis for explaining co-operation is kin selection, whereby individuals accrue fitness benefits by helping close relatives who share many of their genes. Evolutionary game theory has been used to study the evolution of co-operation and kin selection, but until now it has been technically challenging to analyse scenarios where the benefits scale nonlinearly with the number of co-operators, which can occur, e.g., when there are economies of scale in a group activity such as a hunt.
In a new paper led by Nadiah Kristensen, we present a new mathematical method for rigorously accounting for relatedness in evolutionary models with an arbitrary number of players and an arbitrary number of discrete strategies, e.g., co-operators and defectors. We demonstrate the method with an application to a game in which the benefit to individuals rises sharply once some threshold number of co-operators is passed, representing, e.g., a successful hunt. In addition to unconditional co-operators and unconditional defectors, we allow a cognitively advanced strategy: co-ordinated co-operators, who conduct a lottery prior to each game to ensure that the threshold number of co-operators is attained. We find that co-ordinated co-operation is favoured by kin selection. However, if we allow homophily to decline, as happened over the course of human evolution, co-ordinated co-operators can be invaded by another cognitively advanced strategy, the liar, who participates in lotteries but does not follow through on commitments to co-operate. For coordinated cooperation to resist invasion by liars, either some level of homophily must be maintained, or following through on the agreement after a lottery must be in players’ self interest.
Our approach will be broadly useful for exploring the evolution of co-operation in other scenarios involving cognitively advanced strategies that arise from leaps of insight into how the game works. Such scenarios could include, for example, the evolution of punishment and enforcement institutions.
Nadiah was until last year a Research Fellow in our lab, and she is now working at the University of Queensland. This paper was part of an ongoing collaboration with Hisashi Ohtsuki at The Graduate University for Advanced Studies in Japan.
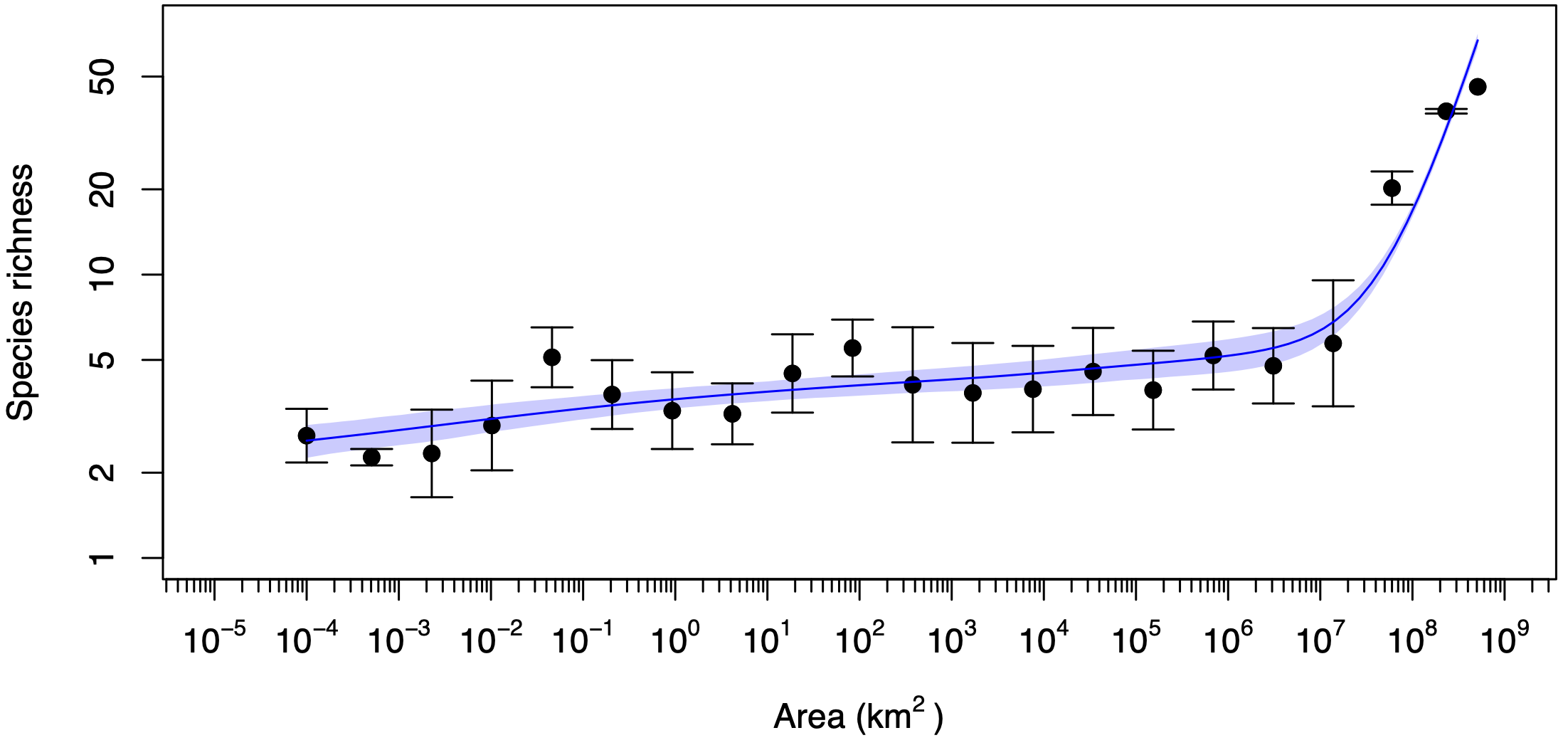
The species–area relationship (SAR) describes how the number of species increases with the area of observation. Classically, the SAR is thought to have three phases: a steep sampling phase at small scales; a shallower power-law phase at intermediate scales; and another a steep phase at very large scales where increases in the logarithm of area reveal new biogeographic realms. More recently, we predicted the presence of a typically hidden fourth SAR phase, sandwiched between the first two classic phases. This elusive SAR phase should be nearly flat, and we call it the niche-structured phase, because within it the number of species is roughly equal to the number of niches. We predicted that the niche-structured SAR phase should be exposed only in unusual scenarios where dispersal or large-scale species richness or both are very low.
In a new paper, just published in Oikos, we discover the elusive fourth SAR phase in a global mangrove dataset. Mangroves are a species-poor group, comprising only around 70 species despite having a global distribution. We found that mangrove species richness exhibits very little change from scales of 100 m2 to 1 million km2, in contrast to a typical power-law SAR, which would exhibit about a factor of 300 increase over this range of scales. Crucially, the species richness did not just collapse to one, which would be a trivial result, but was consistently maintained at about two or three, which we interpret as a measure of the typical number of tidal niches available in a mangrove ecosystem.
This work grew out of the Honours project of NUS undergraduate student Nicholas Fong, and involved a collaboration with mangrove experts Dan Friess and Andre Rovai.
A mangrove tree (Rhizophora spp.) in Peninsular Malaysia. Photo credit: Dan FriessThe species–area relationship for mangroves in the Indo–West Pacific, exhibiting an almost flat phase that spans ten orders of magnitude of area.
Barro Colorado Island (BCI) in central Panama was created when the surrounding forest was flooded in 1914 to create the Panama Canal. The island became a reserve in 1923 and its 15 km2 of tropical forest have been the subject of intensive study ever since. To commemorate 100 years of research on the island, Helene Muller-Landau and Joe Wright, of the Smithsonian Tropical Research Institute, have edited a new two-volume book titled “The First 100 Years of Research on Barro Colorado: Plant and Ecosystem Science“. Ryan has contributed a chapter to this book titled “The Forests of Barro Colorado Island the Neutral Theory of Biodiversity“, which chronicles the history of the neutral theory of biodiversity and how this history has been interwoven with field research at BCI.
Cover art from the book, depicting BCI’s “big tree”, an individual of the species Ceiba pentandra that died in 2013.
We collated forest inventory data at three different scales—local, regional and global—and found signals of synchrony up to scales well beyond 100 km. For the Pasoh and Lambir forests, in particular, there was clear evidence of synchrony for larger trees (greater than 10 cm diameter) although not for smaller trees. A technical challenge in our analyses was developing statistical methods to produce an aggregate estimate of synchrony for each pair of forest sites while accounting for the noise inherent in datasets with large numbers of rare species. Although existing methods are available to estimate synchrony for individual species, these were not appropriate for our forest data because of the short duration of our datasets (a few decades at most) relative to the typical tree generation time. Our novel methods make use of copulas, which are used to model multivariate statistical phenomena in quantitative finance.
We attributed the observed synchrony in tree population dynamics to synchronised environmental drivers, such as climate, although we lacked the statistical power to identify particular drivers. The observed synchrony suggests greater risk of extinction for tree species that are more spatially constrained, especially those with range dimensions less than ~100 km. On the other hand, species with larger ranges may be buffered from extinction by the lack of synchrony in their population dynamics across space.
Forest tree population dynamics are synchronised across space, with synchrony detectable even beyond 100 km. Dark blue, light blue and green curves indicate the results of local, regional and global analyses. Line types indicate the spatial grain of each analysis: synchrony is lower for smaller spatial grains because of immigration.









.jpg){kind=link}
.jpg){kind=link}